Біологія. Довідник школяра та абітурієнта
М’язи як частина опорно-рухової системи
М'язи — органи тіла тварин і людини, які складаються з м’язової тканини, що під впливом нервових імпульсів здатна скорочуватися. М’язи здійснюють переміщення тіла у просторі, зміщення одних його частин відносно інших (динамічна функція), активну фіксацію їх положення відносно одне одного (статична функція), зміну об’єму порожнини тіла або просвітів судин, рухи шкіри та інші функції. У сукупності м’язи утворюють м’язову систему. У людини м’язи становлять від 28-32 % (у жінок) до 35-45 % (у чоловіків) маси тіла.
Залежно від будови м’язових клітин розрізняють гладкі м’язи, які утворюють вісцеральну мускулатуру та попреречно-посмуговані м’язи, що формують парієнтальну мускулатуру. Більшість м’язів, в основному скелетних, відносять до поперечно-посмугованих. М’язи мають різну форму, будову, функцію та розвиток. Переважна більшість кісток скелета сполучається рухомо за допомогою суглобів, завдяки чому утворюються системи важелів. Це пасивна частина системи органів руху та опори організму. Активна частина цієї системи — поперечно-посмуговані м’язи, що за допомогою скорочень і розслаблень змінюють положення кісток, до яких прикріплюються, виконуючи тим самим механічну роботу.
Більшість м’язів прикріплюється до кісток (скелетні м’язи, яких налічують близько 600). Прикріплення їх здійснюється за допомогою сухожилків — тяжів зі щільної волокнистої сполучної тканини, що характерні блискучим білим забарвленням. Лише деякі м’язи функціонують без зв’язку з кістковою системою. Це — колові м’язи, які розташовані, звичайно, навколо природних отворів (рот, задньопрохідний отвір, сечівник) і мають назву стискачів, або сфінктерів.
Особливості будови та функції посмугованих скелетних м’язів
Посмуговані м’язи ще називають скелетними, тому що вони приводять у рух кістковий скелет і є активною частиною опорно-рухового апарату. Завдяки скороченням м’язів виконуються різноманітні рухи, пов’язані з трудовими процесами людини.
Функціональною одиницею посмугованих м’язів є м’язове волокно. Скелетне м’язове волокно має циліндричну форму завдовжки від декількох мікрон до 12,5 см і більше, завтовшки — 10-20 мкм. Саркоплазма скелетного волокна містить мітохондрії, які забезпечують енергетичні процеси діючих м’язів. Крім того, у скелетних волокнах, як і в усіх клітинах, є рибосоми, лізосоми, саркоплазматична сітка та інші органели. У скелетному волокні є специфічні утворення — тоненькі скоротливі волоконця, що отримали назву міофібрили. У кожній міофібрилі під мікроскопом видно темні й світлі диски. В усіх волокнах м’яза як темні, так і світлі диски розташовуються на одному рівні, тому м’язове волокно має вигляд посмугованого.
Темний диск, його ще називають анізотропним (А), розділений світлою смужкою (М), а впоперек світлого диска (І) — ізотропного проходить темна лінія (Т), яка дістала назву телофрагма. Міофібрили не суцільні упродовж скелетного волокна, вони складаються з окремих протофібрил (міофіламентів) завдовжки 1-2 мкм. Крім того, протофібрили неоднорідні, вони є товсті (16 нм) й тонкі (5-7 нм). Товста протофібрила вміщує 180-360 поздовжньо орієнтованих молекул білка міозину, а тонка утворена тоненькими фібрилами іншого білка — актину, який має вигляд подвійної спіралі. Між товстими й тонкими протофібрилами є тоненькі протоплазматичні містки, які утворилися за рахунок білка мероміозину товстої протофібрили. Під час збудження м’язового волокна протоплазматичні містки проштовхують тонкі протофібрили між товстими, за рахунок чого саркомер і загалом усе м’язове волокно вкорочується, м’яз скорочується й приводить у рух кістку, до якої прикріплюється.
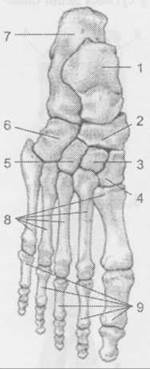
Кістки стопи: 1 — надп’яткова; 2 — човноподібна; 3 — проміжна клиноподібна; 4 — медіальна клиноподібна; 5 — латеральна клиноподібна; 6 — кубоподібна; 7 — п’яткова; 8 — кістки плесна; 9 — фаланги пальців
Тонкі протофібрили, які проникають крізь телофрагму, прикріплюються до неї й далі проходять між товстими міофібрилами.
На межі між дисками А та І сарколема вигинається й утворює Т-трубочки, які розгалужуються всередині волокна. В усіх посмугованих м’язових волокнах добре розвинена незерниста ендоплазматична сітка, яка оточує кожен саркомер. Ці сітки між собою з’єднуються, а їхні канали на межі між саркомерами утворюють кінцеві цистерни, що розташовуються паралельно з Т-трубочками й контактують з ними. Саркоплазма скелетних волокон багата на білок міоглобін, що, як і гемоглобін крові, приєднує Оксиген. Залежно від умісту міоглобіну м’язові волокна бувають червоні й білі. У червоних волокнах міоглобіну та мітохондрій значно більше, ніж у білих. Білі волокна товщі й скорочуються швидше, ніж червоні, а тому вони й стомлюються швидше. У людей скелетні м’язи мають обидва типи волокон, тоді як у м’язах птахів більше червоних волокон.
М’язові волокна містять близько 75 % води й 25 % сухих речовин, до складу яких входять білки, жири, вуглеводи, мінеральні солі й екстрактивні речовини. Кожне волокно має двошарову оболонку (сарколему), під якою в саркоплазмі міститься багато ядер (від кількох до сотень).
Через це волокно за своєю будовою не відповідає особливостям клітинних структур і має назву симпласт. Скорочення скелетних м’язів супроводжується виділенням тепла, за рахунок якого підтримується стала температура тіла. Завдяки скороченням скелетних м’язів поліпшується рух крові та лімфи по кровоносних і лімфатичних судинах, усмоктування їжі у травному тракті та здійснюються всі функції організму.
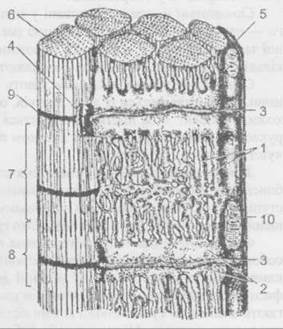
Схема будови відрізка м’язового волокна: 1 — саркоплазматичний ретикулум; 2 — термінальні цистерни саркоплазматичного ретикулуму; 3 — Т-трубки; 4 — тріади; 5 — сарколема; 6 — міофібрили; 7 — А-диск; 8 — І-диск; 9 — Ζ-лінія; 10 — мітохондрії
М’яз як орган
Скелетний м’яз побудований з пучків посмугованих м’язових волокон, укритих пухкою сполучною тканиною, що з’єднує їх між собою. У кожний пучок входить багато м’язових волокон, які утворюють самостійні рухові одиниці. У рухових одиницях може бути від 5-8 до 2 000 і більше м’язових волокон. Кожна рухова одиниця іннервується одним мотонейроном. М’яз складається із черевця й двох сухожилкових кінців. Проксимальний кінець м’яза називають головкою, а дистальний — хвостом. Від головки й хвоста відходять сухожилки, якими м’яз прикріплюється до кісток або фасцій.
Сухожилки — це волокна щільної сполучної тканини. Охоплюючи кінці м’язових волокон, сухожилкові волоконця з’єднуються із саркоплазмою м’яза, що сприяє утворенню міцного зв’язку сухожилка з м’язом. Сухожилки, на відміну від черевця, головки й хвоста, мають білий колір, у них відсутні кровоносні судини, зате багато нервових закінчень та колагенових волокон і мало — еластичних. Через це сухожилки характеризуються великою опірністю до розтягування. У м’язі є пучки волокон різних порядків. Пучки першого порядку, з’єднуючись між собою, утворюють пучки другого, третього та інших порядків. Внутрішню сполучну тканину, яка їх укриває, називають ендомізій, а сполучну тканину, що вкриває м’яз іззовні, отримала назву перимізій.
Крім основних частин м’яза, він має ще й допоміжний апарат: фасції, слизові сумки, сухожилкові зв’язки, піхви, міжм’язові перегородки, фасціальні вузли, утримувачі сухожилків і м’язів, кістково-фіброзні піхви, або фіброзні піхви, блоки.
Фасції. Кожен м’яз або група м’язів (наприклад, група згиначів і розгиначів плеча, передпліччя) оточені тонкою пластинкою зі сполучної (фіброзної) тканини, яку називають фасцією. Фасція відокремлює одну групу м’язів від іншої. Окремі пластинки розгалуженої фасції проникають углиб тіла між м’язами й тому мають назву глибокі. На кінцівках відростки фасцій проникають між групами м’язів і прикріплюються до окістя кісток. Фасції можуть служити місцем прикріплення м’язів, а також своєрідним бар’єром для поширення інфекцій чи запального процесу від одної групи м’язів до іншої. Фасції оточені нервами, кровоносними та лімфатичними судинами, що обслуговують м’язи. Будова фасцій залежить від функції м’язів, їхньої сили. Найкраще розвинені фасції у людей фізичної праці та у спортсменів.
Синовіальні сумки розташовані у місцях прикріплення сухожилків м’язів до кісток (здебільшого — навколо великих суглобів), щоб зменшити тертя кістки з сухожилком. Утворюються зі сполучної тканини, в якій з’являються порожнини з гладенькими стінками. Сумки заповнені невеликою кількістю синовіальної рідини. Розвиваються вони одразу після народження дитини.
Синовіальні піхви сухожилків мають форму циліндричного мішка й утворені зі сполучної тканини. Піхва складається із двох листків, один з яких приростає до сухожилка, а другий охоплює сухожилок ззовні. Між листками міститься синовіальна рідина. Під час скорочення м’яза сухожилок рухається разом з прирощеним листком піхви, а синовіальна рідина зменшує тертя. Такі піхви оточують фаланги кисті, стопи.
Зв’язки являють собою потовщення сполучної тканини, або фасції. Зв’язки міцні й мають вигляд блискучих фіброзних пучків над сухожилками м’язів. Вони утворюють манжету прикріплення до виступів кісток, щоб утримувати у певному положенні сухожилки під час скорочення м’язів. Потовщення, утворені фасціями між окремими групами м’язів, називають міжм’язовими перемичками.
Фасціальні вузли — це потовщення фасцій, розташованих у місці з’єднання двох фасцій між собою. Вони зміцнюють фасціальні піхви судин і нервів. Ці фіброзні утворення зростаються з кістками й доповнюють скелет. У ділянці деяких суглобів (променезап’ястковий, гомілковостопний) фасції потовщуються, утворюючи при цьому утримувачі сухожилків або м’язів. Утримувачі запобігають зміщенню сухожилків і м’язів під час їх скорочення. У розташованих під ними порожнинах, так званих кістково-фіброзних, або фіброзних, піхвах, проходять сухожилки м’язів. В окремих випадках фіброзні піхви спільні для кількох сухожилків.
Блоки локалізуються на епіфізах кісток у вигляді валиків з виїмками, укритих хрящем. На блоках легко рухається, не зміщуючись й не змінюючи напрямку, сухожилок. Крім того, за рахунок блоків збільшується плече волокна й при цьому зростає сила м’яза. Блоками служать сесамоподібні кістки, які є похідними шкіри. Утворюються вони за зміни напрямку сухожилка внаслідок різноманітних рухів, не передбачених морфологічними особливостями наявних утворень. Найбільшою сесамоподібною кісткою є наколінник, значно меншою — горохоподібна кістка та ін.
Механізм скорочення м’яза
М’язове волокно скорочується внаслідок скорочення послідовно з’єднаних саркомерів. За цього довжина актинових і мізинових міофіламентів не змінюється, а зменшується ширина світлих І-дисків. За допомогою світлового мікроскопа й рентгеноструктурного аналізу з’ясовано, що цей процес є наслідком втягування тонких міофіламентів у проміжки товстими за принципом ковзання. Тому теорію, що пояснює процес скорочення м’язів, назають «теорія ковзання». Потенціал дії кінцевої пластинки є причиною виходу медіатора ацетилхоліну, який викликає деполяризацію поверхні сарколеми м’язового волокна у місці розташування кінцевої пластинки. Він розповсюджується і по поверхні сарколеми вздовж волокна, і по перетинках, які вкривають трубочки Т-системи, і всередину волокна, викликаючи деполяризацію перетинок цистерн, яка сприяє швидкому виходу іонів Кальцію з них. Саме іони кальцію запускають процес скорочення. Са2+ зв’язуються із тропоніном (два іони Са2 прикріплюються до однієї молекули тропоніну). Унаслідок цього молекула тропоніну глибше занурюється в жолобки між закрученими нитками актину. Так знімається перешкода контакту між головками міозину й кульками актину, які з’єднуються між собою, утворюючи містки.
Одразу після цього голівка міозину повертається на 45°, розвиваючи певне напруження, яке передається на шийку. За рахунок еластичних властивостей шийки два міофіламенти просуваються один біля одного до центру саркомера.
Такий поворот голівки поперечного містка нагадує рух весла. За одне «веслування» саркомер скорочується лише на 1% своєї довжини. Після того як буде закінчено один акт «веслування», голівка міозину розкриває зв’язок з актиновими міофіламентами і знову з’єднуються під прямим кутом з новою кулькою актину, розташованого ближче до Ζ-пластинки. Таке послідовне з’єднання поперечних містків повторюється багато разів до повного скорочення м’язового волокна. Якщо м’яз скорочується на половину своєї довжини, то в кожному саркомері повинно відбуватися 50 «веслувальних» рухів.
Підвищена концентрація іонів Са2+ у міофібрилярному просторі зберігається всього кілька мілісекунд, а потім кальцієвий насос перекачує Кальцій у цистерни саркоплазматичної сітки. Після того як з молекулярного простору зникне Са2+, тропімінозин знову починає гальмувати міозин-АТФ-азу. Взаємодія поперекових містків припиняється, коли набувають своєї попередньої форми — м’яз розслабляється.
Робота м’язів
Під час будь-якого скорочення м’яз виконує внутрішню роботу, пов’язану з процесами, які відбуваються у м’язовому волокні: рух іонів під час збудження, скорочення й після нього, тертя, втрати енергії у процесі ресинтезу АТФ. Кількісно цю роботу визначають за інтенсивністю споживання Оксигену. Зовнішня (механічна) робота виконується лише під час переміщення будь-якого вантажу, тіла або його частин у просторі.
Механічна робота м’яза (А) вимірюється добутком ваги піднятого вантажу (Р) на вкорочення м’яза (l), тобто: А = Р • l. М’яз має цікаву властивість — за поступового збільшення вантажу його вкорочення за того самого подразнення спочатку зростає, а потім поступово зменшується і за певних значень важкого вантажу дорівнює нулю.
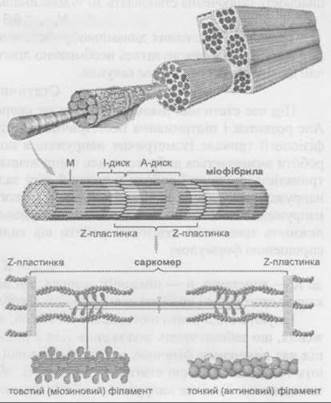
Скоротливий елемент міофібрили
Звідси випливає, що в кожного м’яза максимальна робота можлива лише за певних середніх навантажень. Це — закон середніх навантажень, який випливає із залежності сили скорочення м’яза від його довжини. Відношення зовнішньої роботи м’яза до його внутрішньої роботи називають коефіцієнтом корисної дії (ККД).
Його значення залежить від навантаження й коливається у межах 30-50 %, що можна порівняти з ККД дизельного двигуна (37-41 %).
Треновані м’язи працюють значно економніше, ніж нетреновані, крім того, унаслідок тренування м’язи стають сильнішими й витривалішими. Під час тривалої м’язової роботи відбувається головним чином аеробний ресинтез АТФ за рахунок оксидного фосфорилювання. Необхідна для цього енергія надходить завдяки окисненню вуглеводів і ліпідів. За умови, що швидкість утворення АТФ дорівнює швидкості його розщеплення, система перебуває у рівноважному стані, вміст АТФ і креатинфосфату не змінюється. Під час виконання спортивних вправ, які потребують витривалості, швидкість розщеплення АТФ значно зростає порівняно зі станом спокою, збільшується швидкість ресинтезу АТФ, що можливе за швидкого надходженняу м’яз Оксигену й глюкози. Внаслідок місцевого розширення кровоносних судин швидкість кровотоку в м’язі зростає до 20 разів (робоча гіперемія). Хвилинний об’єм крові, частота скорочень серця й дихання збільшується у 2-3 рази. Перед закінченням тривалої, виснажливої роботи можливий так званий фінішний ривок, зумовлений розщепленням додаткової кількості глікогену анаеробним шляхом за допомогою гліколізу. Під час цієї реакції утворення АТФ відбувається у 2-3 рази швидше, що відповідно забезпечує зростання у 2-3 рази механічної роботи. Щоправда, через обмеження резервів анаеробної енергії граничний час для такої посиленої роботи не перевищує 30 секунд. В умовах інтенсивної м’язової роботи, коли сила скорочення м’язів перевищує 40 % максимальної, внаслідок часткового перетискання кровоносних судин у м’язах відбувається погіршення кровопостачання та зменшення надходження Оксигену. Робота забезпечується за рахунок анаеробного розщеплення креатинофосфату й глікогену, в організмі накопичується значна кількість недоокиснених речовин, зокрема молочної кислоти, виникає киснева заборгованість. Такий борг ліквідується після роботи в результаті саморегуляторного розширення кровоносних судин: у м’язах, що працювали, — реактивна гіперемія.
Діяльність м’язів проявляється у виконанні рухів, пов’язаних з переміщенням тіла або його частин у просторі (динамічна діяльність) і в утриманні тіла або його частин у певному положенні (статична діяльність).
Динамічна діяльність
Динамічна діяльність супроводжується почерговими скороченнями та розслабленнями м’язів та виконанням роботи, яку називають динамічною роботою. Динамічна робота завжди характеризується певною потужністю (N), яка може бути визначена як відношення величини виконуваної роботи (А) до часу її виконання (t), або як добуток сили (F) на швидкість скорочення м’яза (ν), тобто
N = F/t = F • ν.
Експериментально встановлено, що максимальну потужність м’яз розвиває, коли навантаження й швидкість скорочення становлять 30 % максимальної величини, тобто
Nмакс. =0,3 Fмакс. • 0,3 Vмакс.
Тривалість виконання динамічної роботи залежить від її потужності. Динамічна робота невеликої потужності може виконуватись необмежено довго, а за збільшення потужності до максимальних величин динамічна робота триває секунди.
Статична діяльність
Під час статичної діяльності м’язи не скорочуються, тому їх зовнішня робота дорівнює нулю. Але розвиток і підтримання ізометричного напруження потребують певних витрат енергії. Тому у фізіології тривале ізометричне напруження позначають як статичну роботу. Величина статичної роботи визначається добутком сили напруження (F) на тривалість його підтримання (t). Максимальна тривалість статичної роботи у значній мірі залежить від величини напруження. Збільшення сили напруження всього на 10 % призводить до різкого зменшення тривалості статичної роботи. Коли сила напруження досягає 80% максимальної, тривалість статичної роботи становить 15...20 секунд. Залежність тривалості статичної роботи від сили напруження може бути представлена аналітично спрощеною формулою
tмакс. = K / ((F/Fмакс.)n),
де К — константа, n — показник степеня, що дорівнює приблизно 2,5. Статична робота швидко викликає стомлення і не може довго тривати. Особливо статична робота стомлює дітей.
Існують види статичної діяльності м’язів, які можуть продовжуватись доволі довго. Це напруження, що забезпечують положення тіла в полі сили земного тяжіння або підтримання певної пози під час виконання фізичних вправ чи трудової діяльності. Такі тривалі напруження м’язів називають тонічною формою статичної діяльності, або м’язовим тонусом. Тонус м’язів має рефлекторне походження. Тонічне напруження м’язів виникає внаслідок почергової роботи різних рухових одиниць під впливом імпульсів від мотонейронів. Активність мотонейронів підтримується і регулюється імпульсами від пропріорецепторів м’язів та імпульсів від вищих відділів центральної нервової системи. Тонічні напруження перш за все підтримуються завдяки скороченню повільних і спеціальних тонічних одиниць, які, як відомо, можуть перебувати в режимі тетанічного скорочення за невеликої частоти подразнення. Здатність працювати за низького ритму та великих енергетичних ресурсах забезпечують їм можливість перебувати в напруженні тривалий час.
