Загальна біологія. Збірник задач
Генетика популяцій. Закон Харді — Вайнберга
Вид — це основна систематична одиниця, що реально існує в природі, займає певний ареал і є сукупністю споріднених за походженням організмів, які схрещуються між собою з утворенням плодючих організмів, не схрещуються з іншими видами і якісно відмінні від них. Уся сукупність представників будь-якого виду складається з популяцій.
Популяцію можна визначити як сукупність організмів одного виду, що заселяють певну територію, вільно схрещуються між собою і певною мірою ізольовані від інших популяцій даного виду. Це форма існування виду в даних умовах. На основі генетичних змін у популяціях відбуваються мікроеволюційні процеси, які завершуються видоутворенням. Вивчення цих процесів має для теорії селекції та еволюційного вчення першорядне значення.
Вид є генетично закритою системою, популяція — система генетично відкрита. Тому процес видоутворення (у загальній формі) зводиться до перетворення генетично відкритих систем у генетично закриті. Незважаючи на дискретну будову спадкових факторів, мінливість організмів має безперервний характер, а еволюційний процес — тривалим в часі. Популяція і є його елементарною одиницею.
Фундаментальне значення для генетики популяцій мали роботи С. Четверикова. У 1926 році він опублікував статтю «Про деякі моменти еволюційного процесу з точки зору сучасної генетики», у якій обґрунтував вчення про генетичну структуру популяцій, розробив методи генетичного аналізу спадковості популяцій і показав, що всі еволюційні події відбуваються всередині популяції, яка «насичена» мутаціями. Після опублікування цієї роботи Н. Дубінін, С. Райт та інші вчені розгорнули експериментальні й теоретичні дослідження, на основі яких потім сформувалось вчення про генетику популяцій і виникла популяційна генетика. Вона вивчає закони динаміки генетичних перетворень, що відбуваються в природних і експериментальних популяціях тварин, рослин і мікроорганізмів.
У 1908 році англійський математик Г. Харді й німецький лікар Н. Вайнберг незалежно один від одного встановили закон, якому підкоряється частота розподілу гетерозигот у популяції, представники якої вільно схрещуються, і представили його у вигляді алгебраїчної формули. З’ясували, що частота членів пари алельних генів у популяції розподіляється відповідно до коефіцієнта розкладу бінома Ньютона (р + g)2.
Закон Харді — Вайнберга виражає вірогідний розподіл генотипів у популяції, представники якої вільно схрещуються. Але дія цього закону передбачає виконання ряду обов’язкових умов:
- 1) популяція має необмежено велику чисельність;
- 2) всі організми в популяції можуть вільно схрещуватися;
- 3) гомозиготні й гетерозиготні за даною парою алелей особини однаково плодючі, життєздатні і не піддаються добору;
- 4) прямі та зворотні мутації відбуваються (виникають) з однаковою частотою або так рідко, що ними можна знехтувати.
Цілком очевидно, що всі ці умови в реально існуючих популяціях дотримати неможливо, тому закономірності, встановлені Харді та Вайнбергом, діють тільки в ідеальній популяції. Але цей закон є основою для аналізу динаміки генетичних перетворень в реальних природних популяціях у разі порушень, що обумовлені дією еволюційних факторів і добору, виникненням мутацій, обмеженням чисельності популяції тощо. Цей закон необхідний для вивчення будь-яких еволюційних процесів.
Розподіл алелей в необмежено великій популяції за вільного схрещування, відсутності добору і без виникнення мутацій встановлюється на основі концентрації генів, які є в популяції. Концентрація генів — це відносна частота їх в популяції.
Розглянемо співвідношення генотипів у популяції за парою алельних генів А і а. Виразимо частоту гена А величиною р, а частоту його рецесивної алелі а — через g. Оскільки кожний ген однієї алельної пари може бути або А, або а, їхні частоти в сумі дають:
p + g = 1.
Отже, знаючи частоту одного гена, можна визначити частоту іншого. Звідси: якщо частота гена А рівна р, то частота гена а дорівнює 1 - р. У зв’язку з рівномірним розподілом генів між особинами, у них утворюється р яйцеклітин з геном А і g сперміїв з геном а. Оскільки схрещування відбувається вільно, в однаковій мірі можливе поєднання між собою всіх вказаних жіночих і чоловічих гамет.
Можливі комбінації гамет в популяції за вільного схрещування.
|
Спермії |
Яйцеклітини |
|
|
рА |
qa |
|
|
рА qa |
Р2АА РqAа |
pqAa q2aa |
У підсумку одержимо: р2АА + 2gAa + g2aa.
Цей вираз є формулою закону Харді — Вайнберга, з якого випливає, що:
- 1) кількість гомозиготних домінантних особин дорівнює квадрату частоти домінантного гена (р2);
- 2) кількість гомозиготних рецесивних особин дорівнює квадрату частоти рецесивного гена (g2);
- 3) кількість гетерозиготних особин дорівнює подвоєному добутку частот обох алелей (2pg).

За законом Харді — Вайнберга в популяції, представники якої вільно схрещуються, вихідне співвідношення в потомстві гомозигот (домінантних і рецесивних) і гетерозигот залишається постійним. Наприклад, у популяції, у якій розподіляється одна пара алельних генів А і а, особини матимуть один із трьох генотипів: АА, Аа або аа. Інші поєднання цих генів неможливі.
Припустимо, що ці генотипи знаходяться в популяції у співвідношенні 1/4АА + 1/2Аа + 1/4аа. Оскільки можливості для вільного схрещування між собою у носіїв всіх трьох генотипів однакові, то вихідне співвідношення гомозигот і гетерозигот повинно зберігатися в потомстві. Доведемо це, встановивши всі можливі схрещування та їхню частоту.
Після скорочення одержимо: 1/4АА + 1/2Аа + 1/4аа.
Отже, будь-яка популяція, у якій розподіл алельних генів А і а відповідає формулі р2АА + 2pgAa + g2aa, перебуває в стані генетичної рівноваги. Якщо в такій популяції не діє добір і не виникають мутації, вихідна відносна частота алелей зберігається в усіх наступних поколіннях. Використавши формулу Харді — Вайнберга, можна в найпростіших випадках визначити в популяції концентрацію генів певної алельної пари. Якщо в популяції відома частота вияву рецесивного генотипу, то можна обчислити частоту двох інших генотипів даної пари алелей.
Продемонструємо це на прикладі. На ділянці площею 0,25 га нарахували 10 000 проростків кукурудзи. З них 9996 були зеленими, а 4 — жовтими. Відомо, що це явище хлорофільної мутації пов’язане з переходом у гомозиготний стан рецесивного гена. Отже, у цій популяції частота домінантного фенотипу — 99,96%, а рецесивного — 0,04%. Оскільки рецесивний генотип повністю фенотипно виявляється, його частку в популяції можна обчислити:
aа = g2 = 0,0004.
Отже, концентрація рецесивного гена а = √g2 = √0,0004 = 0,02 = 2%. Тоді концентрація (р) домінантного гена А становитиме: 1 - 0,02 = 0,98, або 98%. Тепер можна визначити співвідношення в популяції гомозигот (АА) і гетерозигот (Аа). Кількість гомозигот АА становитиме:
р2 = 0,982 = 0,9604, або 96,04%.
Гетерозиготні рослини Аа мають нормальне зелене забарвлення і ззовні не відрізняються від домінантних гомозиготних рослин АА, хоча несуть рецесивний ген а, який обумовлює утворення жовтих проростків. Визначаємо їх кількість у популяції:
Аа = 2pg = 2 • 0,98 • 0,02 = 0,0392, або 3,92%.
Отже, отримаємо такий розподіл генотипів у популяції кукурудзи:
АА (гомозиготні домінантні, зелені) — р2 = 96,04%;
Аа (гетерозиготні, зелені) — 2pg = 3,92%;
аа (гомозиготні рецесивні, жовті) — g2 = 0,04%.
Фенілкетонурія — небезпечне спадкове захворювання людини, під час якого амінокислота фенілаланін накопичується в крові і виділяється із сечею. Хворі на фенілкетонурію позбавлені однієї з двох фракцій білка, що необхідні для нормального перебігу складної біохімічної реакції перетворення фенілаланіну на тирозин. Встановлено, що фенілкетонурія передається рецесивним геном і виявляється тільки в гомозиготних рецесивних організмів. За даними медичної генетики, концентрація (g) цього гена становить 0,005, а частота гетерозигот — 0,01 (2pg = 2 • 0,995 • 0,005).
Отже, одна людина зі ста є носієм гена фенілкетонурії. Це захворювання супроводжується розумовою відсталістю. Але в разі ранньої діагностики, годування дитини спеціальною дієтою з пониженим вмістом фенілаланіну та регулювання його вмісту в сечі і крові можна помітно поліпшити стан хворого.
Дані про спадкування альбінізму в кукурудзи та фенілкетонурії в людини засвідчують, що, навіть за невеликої частоти небажаних рецесивних гомозигот, кількість гетерозигот, які є носіями цих генів, у популяції відносно велике. При інбридингу, а в людини у випадку споріднених шлюбів, вірогідність появи гомозигот, що мають шкідливі й летальні гени, різко зростає.
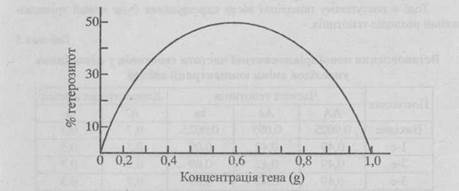
Рис. 1. Залежність розподілу частот генотипів від концентрації алелей (за Лешем)
З формули р2АА + 2pgAa + g2aa випливає, що розподіл генотипів у популяції залежить від концентрації домінантного (р) і рецесивного (g) алельних генів. Залежність розподілу частот генотипів від концентрації алелей досліджена і виражена графічно Д. Лешем.
За даними рис. 1 можна встановити, що помітна зміна частоти гетерозиготних генотипів відбувається тільки у разі значної зміни концентрації алелей. Якщо концентрація алелей коливається в межах 0,35-0,65, відсоток гетерозигот у популяції змінюється несуттєво. У випадку комбінаційної мінливості генотипи можуть розподілятися за двома або кількома незалежними парами алелей. У цих випадках розподіл генотипів відбувається на основі рівновірогідних поєднань компонентів тричленних формул за кожною парою алелей.
Закон Харді — Вайнберга можна назвати законом рівноваги генних концентрацій у популяціях, що вільно схрещуються (панміксичних). Цей закон стверджує, що за відсутності чинників, які змінюють концентрацію генів, популяція може мати будь-які співвідношення алелей і при цьому відносні частоти кожного з них постійні в поколіннях. Якщо популяція з певної причини буде виведена зі стану рівноваги, тобто зміниться відносна концентрація алелей, то в наступному поколінні у ній знову на основі вільного схрещування встановлюється зрівноважений розподіл генотипів. Пояснимо це на такому прикладі (табл. 5).
Вихідні концентрації алелей у популяції становили:
А = р=0,95;
a = g = 0,05.
Рівномірний розподіл генотипів у популяції був таким:
АА = р2 = 0,9025;
Аа = 2pg = 0,0950;
аа = g2 = 0,0025.
Нехай у цій популяції встановились нові концентрації алелей:
А = р = 0,70;
а = g = 0,30.
Тоді в наступному поколінні після схрещування буде новий зрівноважений розподіл генотипів.
Таблиця 5
Встановлення нової зрівноваженої частоти генотипів у поколіннях унаслідок зміни концентрації алелей
|
Покоління |
Частота генотипів |
Концентрація алелей |
|||
|
АА |
Аа |
аа |
А |
а |
|
|
Вихідне |
0,9025 |
0,095 |
0,0025 |
0,7 |
0,3 |
|
1-е |
0,49 |
0,42 |
0,09 |
0,7 |
0,3 |
|
2-е |
0,49 |
0,42 |
0,09 |
0,7 |
0,3 |
|
3-е |
0,49 |
0,42 |
0,09 |
0,7 |
0,3 |
|
n-е |
р2 |
2pg |
g2 |
p |
g |
Ця рівновага генотипів буде зберігатися в популяції доти, доки під впливом якогось чинника знову зміниться концентрація алелей. Схрещування, у результаті якого старий зрівноважений розподіл генотипів перетворюється на новий на основі зміненої концентрації алелей, називають стабілізуючим схрещуванням. Постійна зміна станів рівноваги і перебудова генетичного складу популяції лежать в основі еволюції.
Встановлення в популяції зрівноважених станів генотипів шляхом стабілізуючих схрещувань, згідно закону Харді — Вайнберга, передбачає дотримання наступних умов.
- 1. Концентрація алелей у популяції не повинна змінюватись під впливом прямих і зворотних мутацій. Ген А не змінюється на ген а, і навпаки. Іншими словами, популяція не повинна зазнавати тиску мутаційного процесу.
- 2. Популяція не повинна зазнавати на собі тиску добору. Особини, які мають різні генотипи (АА, Аа і аа), повинні мати однакову життєздатність і плодючість, інакше генетична структура популяції буде змінюватись у результаті добору.
- 3. На популяції не повинен відбиватись тиск міграції, тобто проникнення і входження у схрещування особин з інших популяцій, з іншими співвідношеннями генів.
- 4. Популяція повинна мати настільки велику чисельність, щоб на загальній картині концентрації генів не позначались випадкові відхилення, неминучі під час обмежених вибіркових досліджень.
- 5. На популяцію не повинні впливати чинники ізоляції. Схрещування між особинами повинні здійснюватись на основі випадкових рівновірогідних можливостей всіх генних поєднань популяції.
У реальних (як природних, так і селекційних) популяціях вище названі умови не виконуються. Генетична структура популяцій зазнає безперервних змін: одні генотипи замінюються іншими, у результаті чого в процесі еволюції і селекції змінюється спадковість видів, порід тварин і сортів рослин. Цей процес здійснюється через зміну генетичних структур популяцій під впливом мутацій, природного і штучного доборів, генетико-автоматичних процесів і міграцій.
